As a sensory ecologist, I am interested in how the environment impacts animal behavior and their sensory and communication systems. My research program investigates how behavior and communication are affected when animals and humans meet – specifically when animals come into contact with human-generated noise or urban habitats. How do birds transmit and learn their vocal signals while growing up in noisy areas? On the flip side, can noise be used to keep birds out of areas where they may be pests or become injured, such as agricultural or wind farms?
Previously, in my dissertation research I examined how certain aspects of birdsong may reveal individual quality and are assessed by the individuals they are communicating with – their rivals and prospective mates. Birdsong develops through imitative learning, and the learning environment can impact how those signals are shaped as well as how signals are received.
I addressed the following questions in field and lab settings:
I. Does vocal performance indicate the level of threat to receivers?
II. What factors influence the development of female preferences?
III. How does developmental stress affect adult male vocal performance?
IV. Do males with high vocal performance also possess striking feather colors or do these features trade off?
I. Male vocal performance in the wild: Mating behavior in many species involves communication displays that are vigorous or difficult to perform. Individuals that maximize such features might be of higher quality and thus favored as prospective mates. In songbirds, males produce song both for mate attraction and territory defense. Aspects of vocal performance might allow other males to assess the fighting prowess of signalers in territorial disputes, with males who are better able to sing at higher performance levels perhaps posing a greater threat. Here we tested, in swamp sparrows (Melospiza georgiana), how territorial males respond to playback of songs of various levels of performance. Our main findings are twofold. First, low-performance stimuli elicited significantly lower aggressive responses. Second, male response to normal and high performance songs showed significant individual variation. Closer examination revealed that male response varied with both the subject’s own vocal performance, and the degree to which the performance level of the playback stimuli was elevated. Males approached less closely and spent less time near the speaker the more the high-performance playback had been increased. However, subject males with higher-performance song approached more closely than did males with lower performance song types. These findings provide insight into if and how vocal performance may indicate threat levels posed by a signaler. See Moseley, Lahti, & Podos 2013 PRSB
II. Female mating preferences are a crucial component of sexual selection, yet we have poor knowledge of how female preferences develop. Specifically the extent to which experience during development informs mate choice is largely unstudied. Multiple factors may shape mate choice including experiential learning, social copying, and a sexually selected bias for certain male traits such as the performance level of displays. For bird species in which males learn their songs, it is likely that early exposure to song may also influence females’ preferences later in life. I address this question in the swamp sparrow (Melospiza georgiana), using a new method to elicit preferences from lab-raised birds. Adult, wild-caught females of this species are known to prefer songs of relatively high-performance, i.e. songs that are physically difficult to produce. In 2009, I hand-raised females with tutor songs of normal-performance levels. As further training in their first spring, I presented females with these songs again, but this time coupled with a video of an adult female responding with a copulation solicitation display (CSD, shown below). I then used two methods, the CSD assay and operant conditioning, to test female preferences for songs they had experienced during ontogeny against these same songs altered to higher and lower performance levels. Females gave significantly fewer CSDs to low-performance songs than to the trained (normal-performance) songs. Females responded with the most CSDs to trained songs overall, but this value was not significantly different in a pairwise comparison to the high-performance songs, which was intermediate. A greater response to trained songs supports the hypothesis of a strong influence of early experience, but a preference for high-performance songs by some females cannot be ruled out. In fact in the operant assay, females again elicited both normal and high performance songs significantly more than low performance songs. Thus an interplay of both experiential learning and a bias for high-performance guide female preference development.
III. The development of vocal performance: the effects of nutritional stress, sensorimotor learning, and age. The question of whether animal signals reliably encode information about their bearers is pertinent to investigating how communication signals evolve. The developmental stress hypothesis posits that costs experienced during ontogeny will manifest in signals produced later in life. While developmental stress is known to affect neural attributes involved in song learning, an open area of investigation considers how conditions experienced during ontogeny affect motor performance. Measuring vocal performance, i.e. the ability to produce a difficult signal, is likely to be a fruitful way for receivers to assess individual quality, as executing high-performance signals involves costs and constraints. I hypothesized that developmental stress negatively affects vocal performance, particularly when stress is experienced during both the sensory and sensorimotor phases in the swamp sparrow (Melospiza georgiana). I raised swamp sparrows and trained them with a regime of high performance song models in order to give them a performance challenge during development. I raised these birds in two groups, control and developmental stress conditions; in the latter condition, stressed birds received 70% of the amount of food of control birds during the entire period of hand-feeding. After hand-rearing, supplemental food was restricted and non-edible seeds were mixed in to the seed dispenser both at a two-thirds to one ratio of stress to control. This manipulation continued through the birds’ lives including the sensorimotor phase of song acquisition. Vocal development was recorded until the birds had crystallized song and then again in the birds’ second year of life. Copies made by control birds had significantly higher copying accuracy, consistent with results from a prior study. Additionally, stressed birds produced songs with significantly lower frequency bandwidth, and thus lower vocal performance. Testing all song types with all birds pooled, vocal performance significantly increased between years, with developmentally stressed birds producing songs that had significant increases in frequency bandwidth. These results illustrate previously undocumented negative effects of developmental stress on vocal performance. Variation in birds’ natural environments during sensory and sensorimotor phases may lead to the variation in vocal performance observed in the wild. Thus for receivers, vocal performance may offer a window into assessing birds’ condition early in life. Because vocal performance increased with age, I infer that vocal performance improves with increased practice and could also indicate survival ability. Collaboration with Jeff Podos
IV. Does male vocal performance correlate with other sexually selected traits such as plumage? How does plumage vary between the sexes and with condition? In animal communication, signals operate in diverse modalities. In fact, multimodal signals employing different sensory systems often occur within a single species. Songbirds with extravagant plumage and complex vocalizations are a prime example of species exhibiting flashy traits in multiple modalities. Indeed bird song and color have been the focus of recent literature on this topic (e.g. Uy et al. 2009; O’Loghlen and Rothstein 2009). However, the appropriate aspects of the complex, learned song of oscine birds to measure can be difficult to pinpoint. Ideally one should select song features which are difficult to produce, and thus might signal reliable information (Podos et al. 2010). The swamp sparrow is described as a monomorphic species with the male having a slightly more reddish cap in the breeding season (Pyle 1987). Much is known about the song features of this species in terms of song learning, female preference, and male competition (e.g. Podos 1996, Ballentine et al. 2004; Ballentine et al. 2008). Indeed song features which operate under morphological constraints are well understood in the swamp sparrow (Podos 1996; Podos 1997, Ballentine et al. 2004). Additionally some research has correlated color patches on the head region, specifically the cap, with aggression and paternal care (Olsen et al. 2010). When capturing and banding a population of swamp sparrows in the Quabbin reserve of Massachusetts, I observed striking variation in this color patch both within and between the sexes. This observation prompted questions regarding the degree of sexual dimorphism in this species as well as the capacity for males to be signaling in multiple modes. I examine three main questions regarding the feather color of the cap.
- To what extent is cap color sexually dimorphic: do males and females differ in the spectral qualities and size of their cap feather patches?
- Are color and song signals examples of redundant signals or multiple messages?
- How might cap color and area reliably indicate male quality?
I use the same techniques to measure two key features of the cap color, either of which might be important to visual communication in the swamp sparrow: the intensity of the chestnut color and the proportional area of chestnut relative to black in the cap. For the first feature we use a spectrophotometer to measure reflectance of the chestnut area of the cap. For the second I use the program ImageJ to measure the absolute and relative area of the two head patches – chestnut and black – from high-quality digital images taken on-site in the field or in the lab. Collaboration with David Lahti
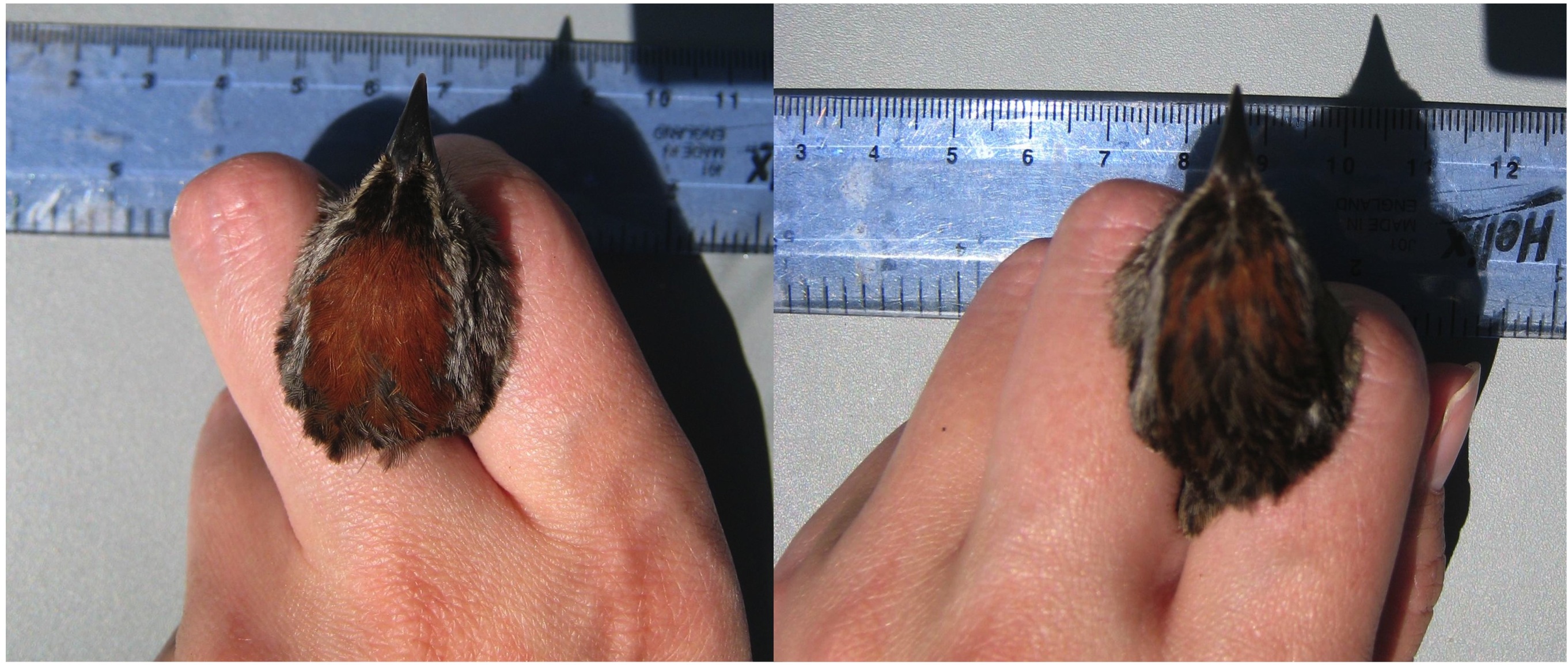
Photo: left – male swamp sparrow, right – female swamp sparrow